Paleoautóctonas (65): Quercus
| Quercus | Familia: Fagaceae | Orden: Fagales | ||
Árboles, menos frecuentemente arbustos. Yemas de ovoideo-oblongas a subglobosas. Hojas pinnatilobadas o pinnatífidas, con menos frecuencia serradas, dentadas o enteras, perennes, marcescentes o caducas; estípulas de alesnadas a linear-espatuladas, ± prontamente caedizas. Flores unisexuales; las masculinas, en amentos laxifloros colgantes, con perianto acampanado o pateniforme, de (4)6(9) lóbulos; estambres por lo general 6-12, de anteras exertas; las femeninas, en amentos laxifloros, en glomérulos o solitarias, axilares, cada una en el interior de un involucro constituido por numerosas escamas, con perianto de acampanado a urceolado; ovario tricarpelar. Frutos (glandes) de maduración anual o bienal, denominados vulgarmente bellotas; aquenios de oblongo-elipsoidales a globosos, 1 por cúpula, de sección redondeada; endocarpo de glabro a tomentoso; cúpula hemisférica o discoidal, entera, coriácea, escamosa, que abarca solamente la base del aquenio; escamas aplicadas y ± concrescentes o en parte libres, las que pueden terminar a veces en puntas blandas o punzantes. do Amaral Franco J. / in: Castroviejo & al. (eds.), Flora iberica vol. 2 / http://www.floraiberica.org / Licencia: Creative Commons |
||||
El género Quercus es uno de los constituyentes más importantes de los bosques templados del Hemisferio Norte y tiene un área de distribución holártica que las glaciaciones apenas han logrado alterar. Esta persistencia del género Quercus en todo el Hemisferio Norte a través de todo el Cuaternario, cuando muchísimos otros géneros desaparecieron de amplísimas zonas, se debe probablemente en gran medida a la extraordinaria capacidad que tienen las especies de este género para migrar y para hibridarse unas con otras.

Hojas de Quercus castaneifolia, especie termófila nativa del Cáucaso y el N de Irán. Arboretum Robert Lenoir, Rendeux, Bélgica / Autor: Jean-Pol GRANDMONT / Licencia: CC BY-SA 3.0
La capacidad de migrar con cierta facilidad se debe al tipo de fruto que produce (bellotas), que es un elemento esencial en la dieta de muchos animales, tanto de mamíferos como aves, que dispersan sus semillas muy eficazmente. Buena prueba de ello es la presencia de este género en las Islas Canarias hasta hace pocos milenios (de Nascimento L. et al., 2009), donde probablemente fue llevado por alguna ave y donde tal vez fue realmente observado el quejigo andaluz (Q. canariensis), cuyo nombre específico se atribuye generalmente a un error de etiquetado (¿Desvelado el misterio del roble canario?).

Expansión de los robles caducifolios en Europa tras la última glaciación. Los círculos coloreados muestran los principales refugios durante la glaciación (Gieseke et al. 2013)
La capacidad de hibridarse y de retrohibridarse ofrece también a los Quercus una manera mucho más sutil de migrar a distancias considerables. En este caso, no son las semillas las que se desplazan, sino el propio genoma del árbol, dispersado por el viento en los granos de polen. Basta con que exista en algún sitio alguna población capaz de hibridarse con otra especie más o menos alejada para que el influjo de genes propicie un desplazamiento de una especie por la otra. Un escenario digno de las mejores películas de ciencia ficción pero bien real.
Aún así, estamos hablando de desplazamientos relativamente lentos a escala de una vida humana, habiendo tardado este género varios milenios en recolonizar Europa tras la última glaciación (ver figura anterior).

Cúpulas de Q. lamellosa, típicas de las especies de la sección Cyclobalanopsis. Yunnan, China. / Fotografía: kristofz / Licencia: CC BY
El origen de este género remonta probablemente al Cretácico Superior, habiéndose atribuido al género Quercus o parataxones afines distintos fósiles encontrados en Norteamérica y el E. de Asia. En Europa el género hace su “aparición” en el Paleoceno, formando parte entonces de una vegetación de tipo subtropical con algunos elementos templados (Barrón et al., 2017).
Tal como muestra el árbol filogenético a continuación, el género Quercus se diversificó muy pronto y ya en el Eoceno se habían individualizado los subgéneros y la mayoría de las secciones que conocemos actualmente. Es interesante notar, por ejemplo, que en aquella época ya había una gran diversidad de especies de este género en regiones septentrionales. En sedimentos del N de Groenlandia datados de 40-42 Ma se han descrito hasta 7 especies pertenecientes a cuatros diferentes secciones (Quercus, Lobatae, Protobalanus e Ilex). Las secciones Quercus, Ilex y Protobalanus no llegarían, sin embargo, a colonizar Europa en aquella época, quedando confinado Protobalanus en Norteamérica hasta nuestros días y apareciendo las secciones Quercus e Ilex mucho más tarde.

Filogenia del género Quercus. Hipp A.L. et al. (2020)
Durante todo el Paleogeno y buena parte de Neogeno, las especies del género Quercus presentes en Europa pertenecieron todas a las secciones Cerris, Lobatae y Cyclobalanopsis, haciendo su aparición la sección Quercus en el Oligoceno e Ilex en el Mioceno. Es interesante notar que la sección Lobatae es hoy en día exclusivamente americana tras desaparecer del continente europeo durante el Mioceno.

Importancia de las distintas secciones del género Quercus en el registro fósil europeo. Barrón (2020)
La persistencia de especies de la sección Cyclobalanopsis hasta el Plioceno en regiones como Portugal y Bulgaria es interesante, siendo hoy en día todas las especies pertenecientes a esta sección originarias del SE asiático. Estas estuvieron probablemente asociadas a la presencia de las últimas laurisilvas del continente europeo, en las que otra fagácea muy próxima a Quercus (Trigonobalanopsis) también estuvo presente.
La llegada de la sección Quercus desde el continente americano se hizo aparentemente en dos oleadas, llegando el antecesor de Q. pontica en el Oligoceno y el del clado de los "roburoides" en el Mioceno, tal vez a través de Asia o directamente a través del puente norteatlántico. Este último clado se diversificó rápidamente a finales del Mioceno y en el Plioceno, dando nacimiento a buena parte de las especies de hojas caducas y marcescentes presentes en continente europeo, siendo absolutamente dominante en el W del mismo. En el E, las especies de la sección Cerris encontraron refugios más duraderos y las especies de ambas secciones conviven allá en amplias zonas.
Los representantes de la sección Ilex son pues los últimos llegados en el continente europeo, habiéndose diferenciado las especies de esta sección a partir de antepasados de origen asiático que llegaron a Europa probablemente a principios del Mioceno, expandiéndose a lo largo del brazo oriental del Tethys. Dos especies esclerófilas muy presentes durante todo el Neógeno en toda Europa pertenecían a este clado (Q. drymeja y Q. mediterranea) y eran constituyentes de bosques de climas húmedos o con veranos húmedos, condiciones similares a las que exigen las actuales especies del Himalaya (Q. baloot y Q. floribunda) que se encuentran en la base del clado que alcanzó el continente europeo.
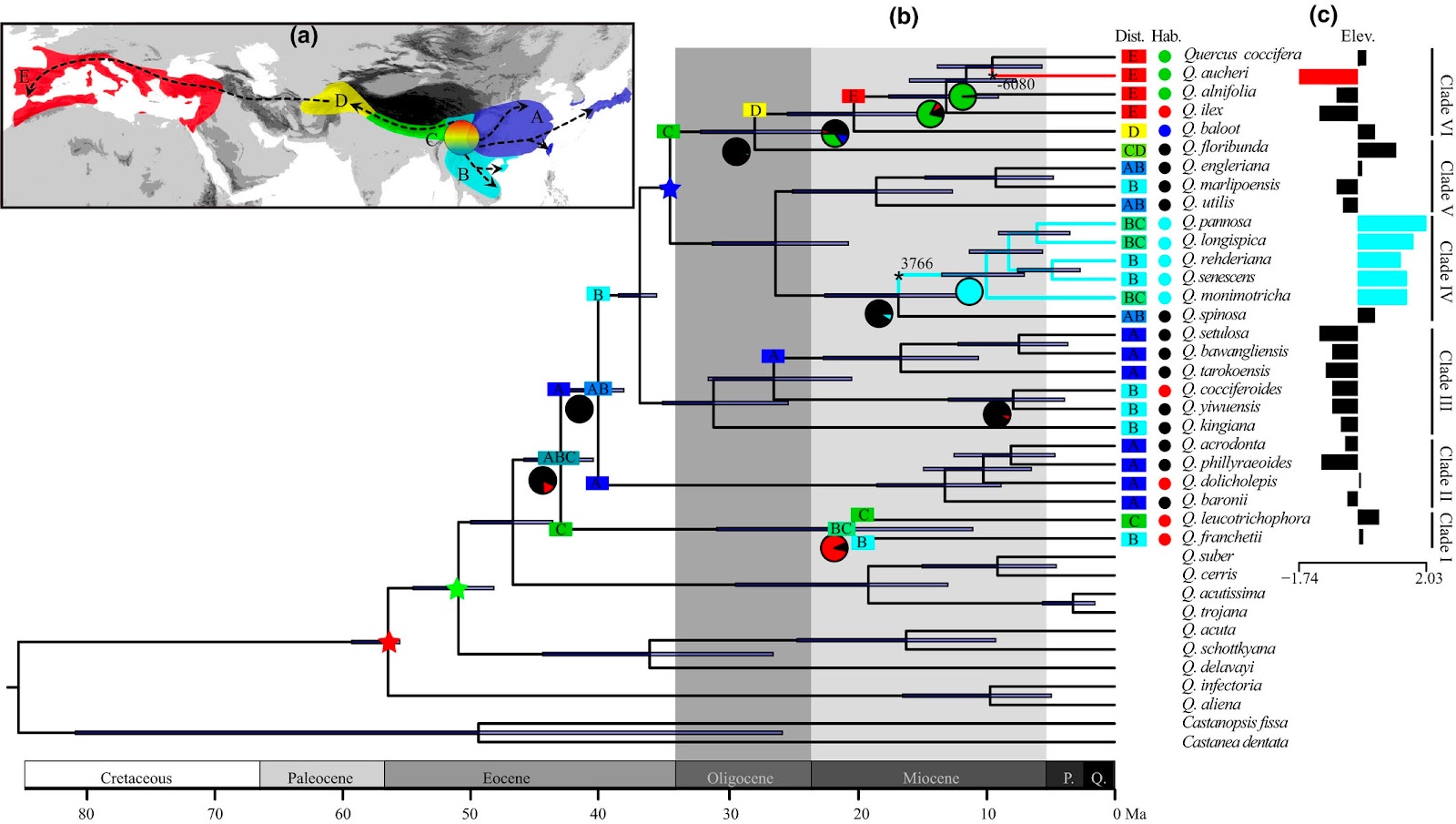
Filogenia de la sección Ilex del género Quercus. Jiang X.L. et al. (2019)
La adaptación a la sequía de las especies de este clado parece haberse iniciado muy tempranamente, habiéndose encontrado polen de tipo ilex-coccifera ya en el Mioceno Medio, por mucho que los macrorestos brillen por su ausencia, probablemente debido al tipo de medios en que se desarrollaba este tipo de vegetación.
Resultado de las idas y venidas del Cuaternario, existe hoy en Europa una buena diversidad de especies adaptadas a climas templados, submediterráneos y mediterráneos. Muy pocas especies, sin embargo, están adaptadas a un clima cálido y húmedo (Cfa según la clasificación de Köppen-Geiger) como el que predominaba en buena parte de Europa antes de las glaciaciones y que los modelos de los climatólogos prevén dominante en toda Europa Central a finales de este siglo. Son fundamentalmente dos las especies de robles termófilas e higrófilas que han sobrevivido en el S del continente europeo o en zonas aledañas (Cólquida, bosque hircano del N de Irán). Una de ellas aún tiene buenas masas en el N de África y sobrevive en algunos puntos de la Península Ibérica. Se trata del quejigo andaluz (Q. canariensis), que según el estudio llevado a cabo por Felicísimo et al (2010), podría ver su área potencial desplazarse hacia el norte de la Península y las zonas más cálidas de Europa Central. Este ejemplo es particularmente interesante porque nos muestra la amplitud de los cambios que hemos de esperar de aquí a finales de siglo.

Distribución potencial del quejigo andaluz (Q. canariensis) en el horizonte 2100 (Felicísimo et al. (2010) / Licencia: Creative Commons).
La otra especie es un poco la contrapartida oriental del quejido andaluz y sus antecesores directos tuvieron una vasta área de distribución en Europa en el Neógeno. El roble persa (Q. castaneifolia) podría, si se confirman los modelos de los climatólogos, hacer un sonado regreso a Europa. No creo, sea dicho de paso, que los ingenieros forestales lo vean con muy mal ojos, al ser esta especie un auténtico monstruo, midiendo algunos ejemplares del N de Irán más de 50 metros de altura con un tronco de más 3,5 m de diámetro.

Este roble persa (Q. castaneifolia), fotografiado en el bosque hircano del N de Irán, mide 60.4 m de altura, 4,9 m de diámetro y 22 m de circunferencia. Es posiblemente el roble más grande del mundo. / Fotografía: Alireza Naqinezhad.
De la sección Cerris, sea dicho de paso, ya se ensaya en muchos países de Europa Central al roble cabelludo (Q. cerris), más resistente a la sequía que Q. robur y Q. petraea, especies con las que convive en Europa Central. No faltan pues alternativas en este género, siempre y cuando se acepte la idea de que el cambio climático obligará muchas especies a buscarse la vida en áreas que les son mucho más favorables. Tratándose de un género tan venerado por los naturalistas, probablemente tardemos un poco en darnos cuenta de la urgencia de la situación, más evidente para aquellas personas que se enfrentan a diario con el problema del decaimiento de muchos de nuestros bosques.

Jiang X.L. et al. (2019) / East Asian origins of European holly oaks (Quercus section Ilex Loudon) via the Tibet-Himalaya / Journal of Biogeography, Vol. 46(10), pp. 2203-2214
Hipp A.L. et al. (2020) / Genomic landscape of the global oak phylogeny / New Phytologist (2020) 226: 1198–1212
Barrón E. et al. (2017) / The Fossil History of Quercus / In: E. Gil-Pelegrín et al. (eds.), Oaks Physiological Ecology. Exploring the Functional Diversity of Genus Quercus L., Tree Physiology 7, https://doi.org/10.1007/978-3-319-69099-5_3
de Nascimento L. et al. (2009) / The long-term ecology of the lost forests of La Laguna, Tenerife (Canary Islands) / Journal of Biogeography, Vol. 36, pp. 499–514
Felicísimo, Á. M. (coord.) 2011. Impactos, vulnerabilidad y adaptación al cambio climático de la biodiversidad española. 2. Flora y vegetación. Oficina Española de Cambio Climático, Ministerio de Medio Ambiente y Medio Rural y Marino. Madrid, 552 pág.









{kind=link}
3 comentarios
De piedra me he quedado con la fotografía del roble persa. Ni idea de su existencia. Gracias.
ResponderEliminarLo mismo me ha pasado a mí, pero se ve que no es un caso único en aquella región...
Eliminarexcelente el estudio, me atrevo a pensar que otro roble como el Quercus frainetto podría añadirse a la lista de robles Cfa por su ecologia y temperamento, otra especie interesante q podría medrar en escenarios futuros a latitudes q hoy ocupan Q robur, petraea o similares
ResponderEliminar